Proper: 3 Models dependents de Amunt: 2 Models independents de Previ: 2.2 Estocasticitat ambiental
Mirem ara una altra font d'estocasticitat que pot produir canvis en el
comportament monòton del model geomètric. Quan ![]() té un valor exactament
igual a
té un valor exactament
igual a ![]() , el nombre de membres de l'espècie es manté constant segons
el model geomètric.
, el nombre de membres de l'espècie es manté constant segons
el model geomètric.
El model geomètric parteix de la idea de que tots els individus es reprodueixen al mateix ritme. Això pot ser cert en organismes molt senzills, però no es compleix en absolut pels organismes superiors.
Quan diem que la natalitat és d'un fill per parella, s'entén que algunes parelles tindran dos, tres o encara més fills mentre que altres no en tindran cap. Cal entendre aquesta afirmació com un promig, no com una indicació de que totes les parelles tenen un fill.
Ambdues coses tan sols són equivalents en el límit en el qual una espècie té infinits membres.
A la pràctica, el nombre de neixements i el de defuncions presenta
fluctuacions. Tant el neixement
d'un ésser viu com la seva defunció
són esdeveniments que no estan exempts d'una certa aleatoritat. La xifra
que ens proporciona el factor de creixement ![]() és tan sols una mitja, la
variància de la qual no és zero.
Aquest fet és especialment notable quan les poblacions són reduïdes.
és tan sols una mitja, la
variància de la qual no és zero.
Aquest fet és especialment notable quan les poblacions són reduïdes.
Intentem plantejar un model que vagi més enllà que el model geomètric en aquest sentit. No podrem plantejar-lo en termes d'una equació de recurrència sinó que caldrà resoldre el model mitjançant una petita simulació.
El model es pot plantejar en termes tan generals com es vulgui, però, per claredat, potser és millor imposar alguna restricció, que en qualsevol cas es pot variar.
Així doncs, per a cada un dels ![]() membres de l'espècie, calculem els
descendents que tenen d'acord amb les següents probabilitats
membres de l'espècie, calculem els
descendents que tenen d'acord amb les següents probabilitats
Per una altra banda, si volem que el nombre de membres de l'espècie es
mantingui constant (per comparar amb el model geomètric ![]() ), la
probabilitat de morir durant un interval de temps ha de ser igual
a la natalitat mitja. En el nostre cas
), la
probabilitat de morir durant un interval de temps ha de ser igual
a la natalitat mitja. En el nostre cas
Un cop simulat el pas d'un interval de temps, en simulem més per poder obtenir l'evolució del nombre de membres d'una espècie.
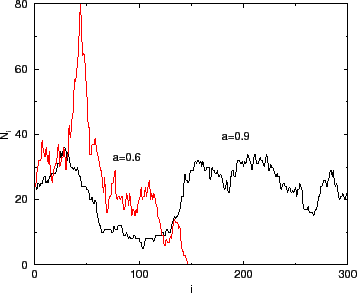
A la Figura 3 es pot observar una simulació del model, partint deAra ja pots fer l'activitat 2.
Taller de simulació medi ambiental